Though there are some theories, the origins of Mycobacterium leprae still remains a mystery. Scientists are not exactly sure how the microbe was able to get into human populations in the first place (Barnes 2005:174). One theory is based on mutation. Mycobacterium vaccae is the closest relative to M. leprae. Some scientists speculate that a more potent variant of M. vaccae could have “gained entry into susceptible individuals and established itself [or mutated into] M. leprae” (ibid:175). If accidentally ingested it may have found it’s way past the submucosal barrier lining of the gut. Making contact with human nerves and skin, it then evolved into a human disease that was transferable via nasal secretions (ibid: 182). This theory, though, is still debated.
Resistance to Drug Treatment In the 1980s, a new technique was used to treat leprosy – multidrug therapy (MDT). “Combination therapy with rifampicin, clofazimine, and dapsone is recommended for the treatment of multibacillary leprosy, whereas therapy with rifampicin and dapsone is recommended for the treatment of paucibacillary leprosy. Ofloxacin plays a key role in the treatment of single-lesion cases and drug-resistant leprosy” (Matsuoka et al 2010:412).
In the 1980s, a new technique was used to treat leprosy – multidrug therapy (MDT). “Combination therapy with rifampicin, clofazimine, and dapsone is recommended for the treatment of multibacillary leprosy, whereas therapy with rifampicin and dapsone is recommended for the treatment of paucibacillary leprosy. Ofloxacin plays a key role in the treatment of single-lesion cases and drug-resistant leprosy” (Matsuoka et al 2010:412).
Many scientists believe that leprosy most likely became viral on the subcontinent of India. Large families were crammed into tight quarters in farming villages with poor living conditions. These situations, we now know, are prime opportunities for leprosy to thrive. But, as history shows, this is clearly not the only way to spread the virus.
Genetics and Pseudogenes
In order to understand the evolution of M. leprae, we must take a look at the different strains of the microbe. Molecular Biologist Stewart Cole of the Pasteur Institute in Paris has found, “M. leprae has the lowest level of genetic diversity of any bacterium I’m aware of […] One clone has infected the whole world” (Grimm 2005:936). His team of scientists have found “subtle DNA sequence mutations called single nucleotide polymorphisms [SNP] that allowed them to break a total of 175 worldwide strains into four types.” These SNPs are broken down to the following areas:
· Type 1 – Asia and the Pacific region
· Type 2 – Ethiopia
· Type 3 – Europe, North Africa, and the Americas
· Type 4 – West Africa and the Caribbean
Comparing the four genome sequences, there is little genomic diversity; they are 99.995% identical. This strengthens the hypothesis that a single clone was passed through an evolutionary bottleneck. “In terms of the diagnosis, treatment and prevention of leprosy, this is extremely encouraging, as it means that antigenic drift in M. leprae should be negligible and the sequences of drug targets will not vary.” (Monot, et al 2009). Cole believes that, by breaking down the mutation patterns among the strains, leprosy most likely originated in either Central Asia or East Africa. He is more inclined to favor leprosy origins in East Africa because “type 2 is the rarest and, thus, likely the oldest.” Furthermore, he feels that “India has been stigmatized as the cradle of leprosy […] but the disease could have just as likely arisen in East Africa” (Grimm 2005:936).
The genome of M. leprae has a very large number of pseudogenes, which means they have lost their protein-coding ability and are thus no longer expressed by the cell. As a result, the rate of growth of the bacillus is extremely slow. This explains why leprosy has such a long incubation period. Additionally, the length of time taken for an infected individual to exhibit symptoms may enhance the ability for the disease to spread. If the individual has undetectable symptoms but is still contagious, the microbes have a greater opportunity to be passed on to unsuspecting victims. (see chart below)


Using pseudogenes to determine the evolution of leprosy, “one group [of scientists] estimated that M. leprae and M. tuberculosis diverged 66 million years ago and that a single pseudogenization event must have occurred in the leprosy bacillus in the last 10–20 million years” (Monot et al 2009). A bulk of the pseudogenes, however, are estimated to have arisen no earlier than 9 million years ago. Undergoing such a drastic change radically altered Mycobacterium leprae. The possibility then exists that M. leprae existed outside of humans before becoming a human pathogen. “It is conceivable that an ancestral form of M. leprae infected an invertebrate host such as an insect, which later acted as a vector for transmitting the bacillus to humans. Support for the latter scenario is provided by studies of the related pathogen Mycobacterium ulcerans, which is at an early stage of reductive evolution and appears to be transmitted to humans by water bugs and/or mosquitoes” (ibid). This still remains a theory; more studies need to be conducted in order to be confirmed.
Regardless of the time and place of origin, the disease has spread with human migration. Trade routes, crusades, colonization, and slavery have all played a pivotal role in making leprosy widespread. But just as important as migration is susceptibility due to an inadequate genetically determined immune response. The incubation period of M. leprae is long and varied. It can range from months to decades, which makes detection extremely difficult. But, because a healthy immune system may be able to ward off the disease, what actually determines if an infected individual will manifest the disease is unknown. Some scientists argue that individual’s genes may be a factor. After the mycobacteria invade the lungs of a healthy individual, phagocytic macrophage-type cells are able to destroy the threatening disease. Even if the immune system does not kill all of the microbes, the disease is not usually fatal. From an epidemiological perspective, this means that leprosy has a greater chance of survival. By killing off an infected host, a disease often minimizes its chances of being passed on. The fact that M. leprae is usually not deadly is beneficial towards its continued existence. But because transmission is difficult, its potential is decreased.


The complexity of leprosy can be further recognized when we analyze the effects M. leprae can have on certain genetic groups. For example, there are some genetically related individuals in India who react to the microbe with localized nerve involvement but without the skin lesions that are typical in most other populations (Barnes 2005:179). Clearly, genetics may determine how susceptible you are to the microbes and how symptoms will be presented if the disease is contracted.
In the 1940s, a new drug called Dapsone was released to combat the spread of the disease. Though it had little effect on the damage already done by leprosy, it was able to control the progression of the disease, and thus prevent further infections. Dapsone was so successful that individuals could be removed from isolation and integrated back into society. They no longer posed a threat to other individuals and as a result, leprosy began to dissipate in many parts of the world.
Dapsone treatments took at least two years to kill all the microbes. In that time social and political outbursts, particularly in many of the endemic regions, prevented proper treatment. With the likes of war and famine health care was often inoperable or out of reach. During these periods, especially if treatment had already begun, the disease had a chance to mutate and develop resistance. M. leprae soon became resistant to the mere treatment of Dapsone (Barnes 2005:183).
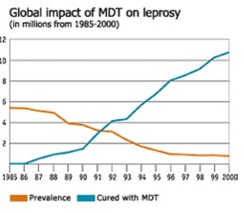 In the 1980s, a new technique was used to treat leprosy – multidrug therapy (MDT). “Combination therapy with rifampicin, clofazimine, and dapsone is recommended for the treatment of multibacillary leprosy, whereas therapy with rifampicin and dapsone is recommended for the treatment of paucibacillary leprosy. Ofloxacin plays a key role in the treatment of single-lesion cases and drug-resistant leprosy” (Matsuoka et al 2010:412).
In the 1980s, a new technique was used to treat leprosy – multidrug therapy (MDT). “Combination therapy with rifampicin, clofazimine, and dapsone is recommended for the treatment of multibacillary leprosy, whereas therapy with rifampicin and dapsone is recommended for the treatment of paucibacillary leprosy. Ofloxacin plays a key role in the treatment of single-lesion cases and drug-resistant leprosy” (Matsuoka et al 2010:412).